



2026. július 17. péntek,
Endre, Elek napja
Endre, Elek napja
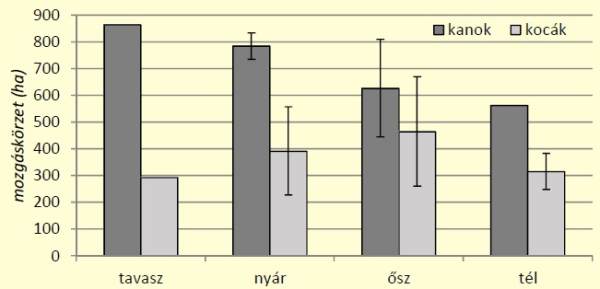 1. ábra: Vaddisznók évszakos mozgáskörzetének alakulása[/caption]
Az eredményeken jól látható, szignifikáns (kétmintás t-próba, p=0,005, n=8), hogy a kanok mozgáskörzete az év minden szakaszában meghaladja a kocákét, ugyanakkor az alakulás tendenciája eltérő. Míg a kanok mozgáskörzete tavasztól (865 ha) télig (563 ha) folyamatos csökkenést mutat, addig a kocák mozgáskörzete tavasszal a legalacsonyabb (293 ha), a nyári növekedést követően ősszel éri el a csúcsot (464ha), majd a tél folyamán újra csökken. A napi aktivitás vizsgálata során kevésbé volt éles a két ivar közötti különbség. A kanok átlagos napi aktivitása éves szinten 4504±703 m volt, míg a kocáké 4300±1312 m. A havi trendet vizsgálva, a két ivar között igazolható különbség nem volt kimutatható (kétmintás t-próba, p=0,638, n=24) (2. ábra).
[caption id="attachment_14900" align="aligncenter" width="600"]
1. ábra: Vaddisznók évszakos mozgáskörzetének alakulása[/caption]
Az eredményeken jól látható, szignifikáns (kétmintás t-próba, p=0,005, n=8), hogy a kanok mozgáskörzete az év minden szakaszában meghaladja a kocákét, ugyanakkor az alakulás tendenciája eltérő. Míg a kanok mozgáskörzete tavasztól (865 ha) télig (563 ha) folyamatos csökkenést mutat, addig a kocák mozgáskörzete tavasszal a legalacsonyabb (293 ha), a nyári növekedést követően ősszel éri el a csúcsot (464ha), majd a tél folyamán újra csökken. A napi aktivitás vizsgálata során kevésbé volt éles a két ivar közötti különbség. A kanok átlagos napi aktivitása éves szinten 4504±703 m volt, míg a kocáké 4300±1312 m. A havi trendet vizsgálva, a két ivar között igazolható különbség nem volt kimutatható (kétmintás t-próba, p=0,638, n=24) (2. ábra).
[caption id="attachment_14900" align="aligncenter" width="600"]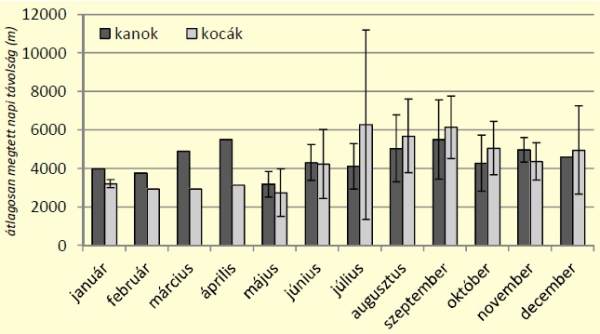 2. ábra: Vaddisznók havi aktivitásának alakulása az átlagosan megtett napi távolságok alapján[/caption]
Az óránkénti aktivitás vizsgálata során a két ivar aktivitási görbéjének lefutása hasonló, mindkettőn két csúcs található. Ezek közül az esti magasabb, míg a hajnali alacsonyabb értéket mutat. A két görbe között mutatkozó eltérést – kanok alacsonyabb napközbeni és magasabb éjszakai aktivitása – nem sikerült statisztikailag igazolni (kétmintás t-próba, p=0,969). Ugyanakkor megállapítható volt, hogy mindkét ivar esetén a legalacsonyabb aktivitási értékkel a 11:00-13:00 közötti időszak rendelkezett (3. ábra).
[caption id="attachment_14899" align="aligncenter" width="600"]
2. ábra: Vaddisznók havi aktivitásának alakulása az átlagosan megtett napi távolságok alapján[/caption]
Az óránkénti aktivitás vizsgálata során a két ivar aktivitási görbéjének lefutása hasonló, mindkettőn két csúcs található. Ezek közül az esti magasabb, míg a hajnali alacsonyabb értéket mutat. A két görbe között mutatkozó eltérést – kanok alacsonyabb napközbeni és magasabb éjszakai aktivitása – nem sikerült statisztikailag igazolni (kétmintás t-próba, p=0,969). Ugyanakkor megállapítható volt, hogy mindkét ivar esetén a legalacsonyabb aktivitási értékkel a 11:00-13:00 közötti időszak rendelkezett (3. ábra).
[caption id="attachment_14899" align="aligncenter" width="600"]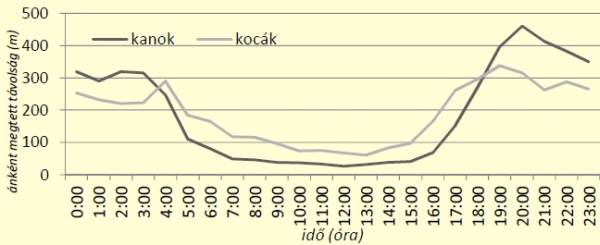 3. ábra: Vaddisznók napi aktivitásának alakulása az átlagosan megtett óránkénti távolságok alapján[/caption]
Az aktivitás alakulását számos tényező befolyásolhatja, ezek közül egyik a társas nagyvad vadászatok. Ezek hatásának vizsgálatához beszereztük a hajtástérképeket, majd azonosítottuk azokat az eseteket, amelyek során a hajtásokban tartózkodott jelölt egyed. Ilyen esetre 4 alkalommal került sor. Meghatároztuk a hajtás napján 8:00 és 16:00 között megtett távolság mértékét, valamint a hajtást megelőző- és az azt követő két-két nap azonos periódusra vonatkozó érékeit (4. ábra).
[caption id="attachment_14898" align="aligncenter" width="600"]
3. ábra: Vaddisznók napi aktivitásának alakulása az átlagosan megtett óránkénti távolságok alapján[/caption]
Az aktivitás alakulását számos tényező befolyásolhatja, ezek közül egyik a társas nagyvad vadászatok. Ezek hatásának vizsgálatához beszereztük a hajtástérképeket, majd azonosítottuk azokat az eseteket, amelyek során a hajtásokban tartózkodott jelölt egyed. Ilyen esetre 4 alkalommal került sor. Meghatároztuk a hajtás napján 8:00 és 16:00 között megtett távolság mértékét, valamint a hajtást megelőző- és az azt követő két-két nap azonos periódusra vonatkozó érékeit (4. ábra).
[caption id="attachment_14898" align="aligncenter" width="600"]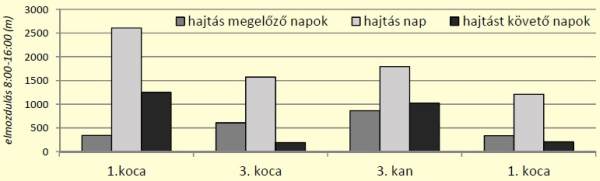 4. ábra: Társas vadászatok hatása a vaddisznó aktivitására[/caption]
Az eredmények alapján elmondható, hogy a hajtás napján a jelölt egyedek minden esetben nagyobb elmozdulásokat produkáltak, mint a szomszédos napokon. A zavart területről történő elmozdulás a hajtás napján az aktivitás alakulásában az előző napokhoz (540±216 m) képest átlagban 3,9-szeres emelkedést (1798±512 m) jelentett, majd a hajtást követő napokon az aktivitás 4,1 szeres- csökkenést mutatott (671±474 m). A különbség szignifikáns volt (párosított t-próba), mind a hajtás megelőző napok és a hajtásnap (p=0,03, n=8) között, mind pedig a hajtást követő napok és a hajtás nap között (p=0,004, n=8). Különbség a hajtást megelőző és az azt követő napok aktivitása között nem volt kimutatható (p=0,674, n=8).
A jelölt egyedek élőhely-használatának és élőhely-preferenciájának vizsgálatához első lépésként meghatároztuk ivaronként az egyes erdőállomány-típusokban található pozíciók számát, majd ezeket %-os értékben adtuk meg (5. ábra).
[caption id="attachment_14897" align="aligncenter" width="600"]
4. ábra: Társas vadászatok hatása a vaddisznó aktivitására[/caption]
Az eredmények alapján elmondható, hogy a hajtás napján a jelölt egyedek minden esetben nagyobb elmozdulásokat produkáltak, mint a szomszédos napokon. A zavart területről történő elmozdulás a hajtás napján az aktivitás alakulásában az előző napokhoz (540±216 m) képest átlagban 3,9-szeres emelkedést (1798±512 m) jelentett, majd a hajtást követő napokon az aktivitás 4,1 szeres- csökkenést mutatott (671±474 m). A különbség szignifikáns volt (párosított t-próba), mind a hajtás megelőző napok és a hajtásnap (p=0,03, n=8) között, mind pedig a hajtást követő napok és a hajtás nap között (p=0,004, n=8). Különbség a hajtást megelőző és az azt követő napok aktivitása között nem volt kimutatható (p=0,674, n=8).
A jelölt egyedek élőhely-használatának és élőhely-preferenciájának vizsgálatához első lépésként meghatároztuk ivaronként az egyes erdőállomány-típusokban található pozíciók számát, majd ezeket %-os értékben adtuk meg (5. ábra).
[caption id="attachment_14897" align="aligncenter" width="600"]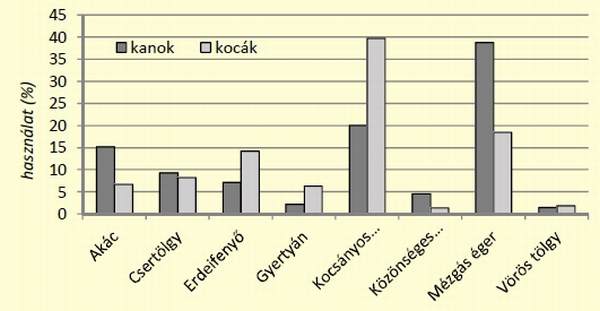 5. ábra: Vaddisznók élőhely-használatának alakulása ivar szerinti bontásban[/caption]
Az ábrán azon állományok szerepelnek, melyek használata meghaladta az 1%-ot. A vizsgált főfafajok közül a kocsánytalan tölgyesek és az égeresek használata haladta meg a 15%-os értéket, a többi fafaj ettől elmaradt. A két ivar között különbség volt. Míg a kanok esetében az éger használata volt a leggyakoribb (38,8%), addig a kocák a kocsányos tölgyet használták magasabb arányban (39,7%).
A használathoz képest az egyes állományok kedveltsége más képet mutatott. A kocák az akác, erdeifenyő, kocsányos tölgy állományokat preferálták, míg a kanok esetében a kedvelt fajok közé tartozott az akác, csertölgy, közönséges nyír és az éger. Az erdőállományok használatának vizsgálata során a két ivar között különbséget nem sikerült igazolni (két mintás t-próba, p=0,528, n=16), ezért a következőkben a legmagasabb használatot mutató két főfafaj (kocsányos tölgy és éger) esetében az elemzéseket ivari bontás nélkül végeztük el. A két állomány használatának havi alakulásának trendjében eltérések figyelhetőek meg. A kocsányos tölgy használata januártól-májusig növekszik, majd csökkenést mutat augusztusig. Ekkor éri el a legalacsonyabb szintet 16,4%-kal, ezt követően újra emelkedés figyelhető meg egészen decemberig, amikor eléri a legmagasabb használatot (44,1%). Az égeresek használata tölgyhöz viszonyítva eltérő képet mutat, januártól-májusig alapvetően alacsony használat figyelhető meg, elérve márciusban a legalacsonyabb értéket (13,1%).
Majd júniusban megemelkedik a használat és szeptemberig egyenletesen magas marad (31-36%), ezt követően mutatkozik csökkenés decemberig bezárólag. (6. ábra). A két adatsor – kocsányos tölgy és mézgás éger használata – elemzésekor statisztikailag igazolható eltérés nem volt kimutatható (párosított tpróba, p=0,378, n=24), ez annak köszönhető, hogy a két állomány használata éves szinten hasonló képet mutat, ugyanakkor annak időbeni változásai különbözőek.
[caption id="attachment_14896" align="aligncenter" width="600"]
5. ábra: Vaddisznók élőhely-használatának alakulása ivar szerinti bontásban[/caption]
Az ábrán azon állományok szerepelnek, melyek használata meghaladta az 1%-ot. A vizsgált főfafajok közül a kocsánytalan tölgyesek és az égeresek használata haladta meg a 15%-os értéket, a többi fafaj ettől elmaradt. A két ivar között különbség volt. Míg a kanok esetében az éger használata volt a leggyakoribb (38,8%), addig a kocák a kocsányos tölgyet használták magasabb arányban (39,7%).
A használathoz képest az egyes állományok kedveltsége más képet mutatott. A kocák az akác, erdeifenyő, kocsányos tölgy állományokat preferálták, míg a kanok esetében a kedvelt fajok közé tartozott az akác, csertölgy, közönséges nyír és az éger. Az erdőállományok használatának vizsgálata során a két ivar között különbséget nem sikerült igazolni (két mintás t-próba, p=0,528, n=16), ezért a következőkben a legmagasabb használatot mutató két főfafaj (kocsányos tölgy és éger) esetében az elemzéseket ivari bontás nélkül végeztük el. A két állomány használatának havi alakulásának trendjében eltérések figyelhetőek meg. A kocsányos tölgy használata januártól-májusig növekszik, majd csökkenést mutat augusztusig. Ekkor éri el a legalacsonyabb szintet 16,4%-kal, ezt követően újra emelkedés figyelhető meg egészen decemberig, amikor eléri a legmagasabb használatot (44,1%). Az égeresek használata tölgyhöz viszonyítva eltérő képet mutat, januártól-májusig alapvetően alacsony használat figyelhető meg, elérve márciusban a legalacsonyabb értéket (13,1%).
Majd júniusban megemelkedik a használat és szeptemberig egyenletesen magas marad (31-36%), ezt követően mutatkozik csökkenés decemberig bezárólag. (6. ábra). A két adatsor – kocsányos tölgy és mézgás éger használata – elemzésekor statisztikailag igazolható eltérés nem volt kimutatható (párosított tpróba, p=0,378, n=24), ez annak köszönhető, hogy a két állomány használata éves szinten hasonló képet mutat, ugyanakkor annak időbeni változásai különbözőek.
[caption id="attachment_14896" align="aligncenter" width="600"]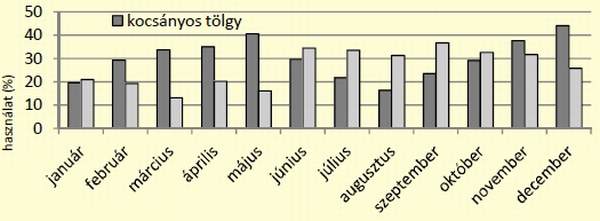 6. ábra: Kocsányos tölgy és mézgás éger állományok használatának alakulása az év során[/caption]
A használat mellett vizsgáltuk a két állomány kedveltségét is. A tölgy esetében mindösszesen három esetben: május, november és december sikerült preferenciát kimutatni, míg az éger esetében az állomány kedveltsége júniustól-decemberig folyamatos volt, szeptembertől csökkenő értékkel (7. ábra).
[caption id="attachment_14895" align="aligncenter" width="600"]
6. ábra: Kocsányos tölgy és mézgás éger állományok használatának alakulása az év során[/caption]
A használat mellett vizsgáltuk a két állomány kedveltségét is. A tölgy esetében mindösszesen három esetben: május, november és december sikerült preferenciát kimutatni, míg az éger esetében az állomány kedveltsége júniustól-decemberig folyamatos volt, szeptembertől csökkenő értékkel (7. ábra).
[caption id="attachment_14895" align="aligncenter" width="600"]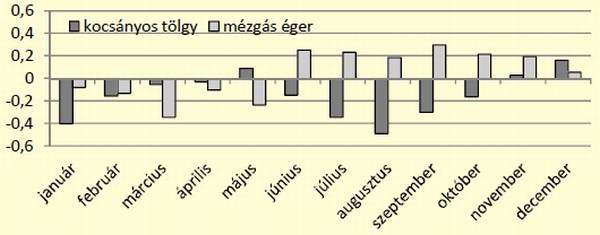 7. ábra: Kocsányos tölgy és mézgás éger állományok preferenciájának alakulása az év során[/caption]
Vizsgáltuk továbbá e két állomány beállóhelyként való használatát is, a nyugalmi időszak pozícióit felhasználva. Eszerint a jelölt egyedek az esetek 38%-ban kocsánytalan tölgyben, 30%-ban pedig égerben töltötték a nappalokat. A részletesebb kép elérése érdekében elkészítettük mindkét fafaj esetében beállóhelyek használatának korosztályi bontását. Eszerint a tölgy esetében a 0-30 éves korosztályt használta, azon belül is a 10-20 év körüli korosztály volt jellemezhető a legmagasabb (41%) értékkel. Az éger esetében két korosztály esetén volt magasabb a használat 10-20 év (28%) valamint az 50-60 év (31%) (8. ábra).
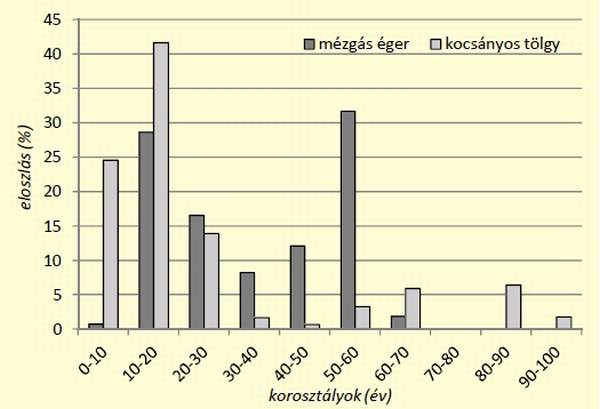
[caption id="attachment_14894" align="aligncenter" width="600"] 8. ábra: Beállóhelyek erdőállományainak korosztályi eloszlása mézgás éger és kocsányos tölgy esetében[/caption]
Eredmények értékelése és következtetések
A mozgáskörzetek meghatározása során szerzett információk alapján elmondható, hogy a kanok nagyobb területet járnak be az év során, mint a kocák, ennek oka feltételezhetően a kocák utódnevelésben betöltött szerepének köszönhető. Ezt támasztják alá a szezonális mozgáskörzet alakulások is, tavasszal rendelkeztek a kocák a legalacsonyabb mozgáskörzettel, ez a periódus egybe esik a malacnevelés kezdeti időszakával (MASSEI ET AL. 1997).
Az érték ezt követően folyamatosan emelkedett az ősz végéig, ez betudható a malacok korosbodásának, illetve a mezőgazdasági kultúrák szezonális változásának és egyben a nyílt élőhelyek intenzívebb használatának. A mozgáskörzetek méretének növekedése azt is jelenti, hogy a területen üzemeltetett etetők-szórók hatékonysága is változhat az év során.
A nem megfelelően megválasztott etetőhálózat problémákat generálhat, mivel túl sűrű hálózat esetén a takarmányozási költségek magasak lehetnek anélkül hogy nőne a takarmányozás hatékonysága. Túl ritkán elhelyezett etetőkkel-szórókkal pedig nem érhető el hatékony takarmányozás. Télen és tavasszal, amikor a kocák a legkisebb mozgáskörzettel rendelkeznek, szükséges lehet az etetők (hatékony takarmányozás) és a szórók (eredményes vadászat) sűrűségének emelése, majd az idő előrehaladtával számuk csökkenthető. A mozgáskörzetek szezonális vizsgálata során kapott eredményeink alapján, a tél és tavasz folyamán optimálisnak tartanánk a 300 ha-onkénti 1 etető üzemeltetését, ez az érték a nyár és ősz folyamán 1etető/500 ha-ra változtatható. Mivel a kanok nagyobb területeket járnak be, a kocák igényeire optimalizált etető elhelyezések megfelelőek számukra is. A két ivar napi aktivitási értékei közel azonosak voltak vizsgálatunk során, különbség a havi értékek trendjében, különösen a malacozásmalacnevelés időszakában voltak megfigyelhetőek. A júniustól –szeptemberig terjedő időszakban, mindkét ivar aktivitása emelkedett. Ebben szerepet játszhattak a mezőgazdasági területekre történő kiváltások, az ekkor megtett távolságok mértékét befolyásolhatta a mezőgazdasági élőhely nyújtotta kínálat térbeli alakulása (NÁHLIK ET AL., 2013). Az óránkénti aktivitás esetében nem volt szignifikáns eltérés a két ivar között. Mindkettő esetében kétcsúcsú aktivitási görbe volt jellemző, amelyhez hasonló aktivitást írnak le más vizsgálatok eredményei is (KEULING ET AL., 2008). Az esti aktivitási csúcsot egy magasabb, de rövidebb ideig tartó elmozdulási érték jellemezte, míg a hajnali egy kisebb értékkel volt jellemezhető, de elnyújtottabb volt az időszak. A két csúcsot éjszaka összekötő időben az aktivitás jócskán meghaladja a nappali, beálló helyen való tartózkodást jellemző kis értéket, amelynek minimuma 11:00-13:00 között van. A görbék lefutásából feltételezhető, hogy a beálló helyről történő kimozdulás a lehető leggyorsabban elérhető táplálék felé irányul, majd onnan folyamatos mozgás mellett további táplálékkeresés illetve táplálkozás történik, ezután a hajnali órákban megnövekedett aktivitás mellett keresi meg a megfelelő beálló helyet. A nyugalmi időszakában történő zavaró hatásra a vaddisznó megnövekedett aktivitással válaszol (MAILLARD AND FOURNIER. 1995). Ezt támasztják alá eredményeink, amelyeket a társasvadászatok hatásának vizsgálatakor nyertünk, a zavaró hatásra a területről a vaddisznó elmozdult, reakciója rövid ideig tartott. A vadászat zavaró hatása azon a napon érvényesült csak, az azt követő időszak aktivitása megegyezett a vadászatot megelőző napok értékével.
7. ábra: Kocsányos tölgy és mézgás éger állományok preferenciájának alakulása az év során[/caption]
Vizsgáltuk továbbá e két állomány beállóhelyként való használatát is, a nyugalmi időszak pozícióit felhasználva. Eszerint a jelölt egyedek az esetek 38%-ban kocsánytalan tölgyben, 30%-ban pedig égerben töltötték a nappalokat. A részletesebb kép elérése érdekében elkészítettük mindkét fafaj esetében beállóhelyek használatának korosztályi bontását. Eszerint a tölgy esetében a 0-30 éves korosztályt használta, azon belül is a 10-20 év körüli korosztály volt jellemezhető a legmagasabb (41%) értékkel. Az éger esetében két korosztály esetén volt magasabb a használat 10-20 év (28%) valamint az 50-60 év (31%) (8. ábra).
[caption id="attachment_14894" align="aligncenter" width="600"] 8. ábra: Beállóhelyek erdőállományainak korosztályi eloszlása mézgás éger és kocsányos tölgy esetében[/caption]
Eredmények értékelése és következtetések
A mozgáskörzetek meghatározása során szerzett információk alapján elmondható, hogy a kanok nagyobb területet járnak be az év során, mint a kocák, ennek oka feltételezhetően a kocák utódnevelésben betöltött szerepének köszönhető. Ezt támasztják alá a szezonális mozgáskörzet alakulások is, tavasszal rendelkeztek a kocák a legalacsonyabb mozgáskörzettel, ez a periódus egybe esik a malacnevelés kezdeti időszakával (MASSEI ET AL. 1997).
Az érték ezt követően folyamatosan emelkedett az ősz végéig, ez betudható a malacok korosbodásának, illetve a mezőgazdasági kultúrák szezonális változásának és egyben a nyílt élőhelyek intenzívebb használatának. A mozgáskörzetek méretének növekedése azt is jelenti, hogy a területen üzemeltetett etetők-szórók hatékonysága is változhat az év során.
A nem megfelelően megválasztott etetőhálózat problémákat generálhat, mivel túl sűrű hálózat esetén a takarmányozási költségek magasak lehetnek anélkül hogy nőne a takarmányozás hatékonysága. Túl ritkán elhelyezett etetőkkel-szórókkal pedig nem érhető el hatékony takarmányozás. Télen és tavasszal, amikor a kocák a legkisebb mozgáskörzettel rendelkeznek, szükséges lehet az etetők (hatékony takarmányozás) és a szórók (eredményes vadászat) sűrűségének emelése, majd az idő előrehaladtával számuk csökkenthető. A mozgáskörzetek szezonális vizsgálata során kapott eredményeink alapján, a tél és tavasz folyamán optimálisnak tartanánk a 300 ha-onkénti 1 etető üzemeltetését, ez az érték a nyár és ősz folyamán 1etető/500 ha-ra változtatható. Mivel a kanok nagyobb területeket járnak be, a kocák igényeire optimalizált etető elhelyezések megfelelőek számukra is. A két ivar napi aktivitási értékei közel azonosak voltak vizsgálatunk során, különbség a havi értékek trendjében, különösen a malacozásmalacnevelés időszakában voltak megfigyelhetőek. A júniustól –szeptemberig terjedő időszakban, mindkét ivar aktivitása emelkedett. Ebben szerepet játszhattak a mezőgazdasági területekre történő kiváltások, az ekkor megtett távolságok mértékét befolyásolhatta a mezőgazdasági élőhely nyújtotta kínálat térbeli alakulása (NÁHLIK ET AL., 2013). Az óránkénti aktivitás esetében nem volt szignifikáns eltérés a két ivar között. Mindkettő esetében kétcsúcsú aktivitási görbe volt jellemző, amelyhez hasonló aktivitást írnak le más vizsgálatok eredményei is (KEULING ET AL., 2008). Az esti aktivitási csúcsot egy magasabb, de rövidebb ideig tartó elmozdulási érték jellemezte, míg a hajnali egy kisebb értékkel volt jellemezhető, de elnyújtottabb volt az időszak. A két csúcsot éjszaka összekötő időben az aktivitás jócskán meghaladja a nappali, beálló helyen való tartózkodást jellemző kis értéket, amelynek minimuma 11:00-13:00 között van. A görbék lefutásából feltételezhető, hogy a beálló helyről történő kimozdulás a lehető leggyorsabban elérhető táplálék felé irányul, majd onnan folyamatos mozgás mellett további táplálékkeresés illetve táplálkozás történik, ezután a hajnali órákban megnövekedett aktivitás mellett keresi meg a megfelelő beálló helyet. A nyugalmi időszakában történő zavaró hatásra a vaddisznó megnövekedett aktivitással válaszol (MAILLARD AND FOURNIER. 1995). Ezt támasztják alá eredményeink, amelyeket a társasvadászatok hatásának vizsgálatakor nyertünk, a zavaró hatásra a területről a vaddisznó elmozdult, reakciója rövid ideig tartott. A vadászat zavaró hatása azon a napon érvényesült csak, az azt követő időszak aktivitása megegyezett a vadászatot megelőző napok értékével.
 Hasonlóan megnövelt aktivitással reagált a gímszarvas a nem állandóan fennálló, intenzív zavaró hatásokra (NÁHLIK ET AL., 2014). Az erdei élőhely-használat vizsgálata során tapasztaltak a vaddisznó kiváló alkalmazkodó képességét támasztják alá (D’HUART, 1991).
Élőhely-használata során törekedett a számára legoptimálisabb faállományok kiválasztására, amelyek táplálkozás és búvóhely szempontjából is megfelelőek. A vizsgálati területen két fafaj állományait használta intenzíven, ezek a kocsányos tölgy és a mézgás éger voltak. A használat havi alakulása jól szemlélteti vaddisznó arra adott reakcióját, hogy a két fafaj állományai által nyújtott táplálék- és búvóhely kínálatban változások történnek. A mézgás éger, magas aljnövényzetével, elsősorban nappali búvóhelyet nyújt a vaddisznó számára, ezt támasztják alá a beállóhelyek használatának elemzésekor szerzett eredményeink. A nedves-vizes élőhely jó dagonyázási lehetőséget jelent, illetve táplálkozó helyként is szerepe lehet, a magas puhatestű táplálék kínálatból kifolyólag, ami kedvelt tápláléka a vaddisznónak (HOWE ET AL. 1981).
A fiatal tölgy állományok ugyanakkor búvóhelyet jelentenek, míg az idősebb állományok makktermésük révén töltenek be fontos szerepet a vaddisznó táplálkozásában, mivel a természetes táplálékok közül ez a legpreferáltabb forrás (FOURNIER-CHAMBRILLON ET AL., 1995). Vizsgálatunkban a tölgy állományok legalacsonyabb használati értéke augusztusban volt megfigyelhető, majd folyamatos emelkedés mellett, decemberben érte el a csúcsot, ezzel párhuzamosan viszont csökkent az égeresek használata. Az augusztus december periódus kezdetén a mezőgazdasági kultúrák nyújtotta táplálék kedveltsége (HERRERO ET AL. 2006) és az égeresek dús cserjeszintje okozhatja a tölgyek alacsony használatát. Majd az ősz során az égeresek takarásának csökkenésével és a mezőgazdasági kultúrák visszaszorulásával párhuzamosan növekedni kezd a tölgyesek használata. A magas novemberi-decemberi tölgy használatban elsősorban a táplálék kínálat játszhat szerepet, ami januárra visszaesik. Ez köszönhető a makk fogyásának, a hótakarónak és a fagyott talajnak, ugyan is a fagyott föld miatt csökken a földből felvett táplálékok aránya a táplálékban (HERRERO ET. AL, 2005). Az égeresek használatának márciusig folytatódó csökkenésével a tölgyesek használata emelkedik, ez az égeresek nyújtotta takarás további csökkenésével hozható összefüggésbe. A vegetáció megindulásával áprilistól kezdődően mind az éger, mind pedig a tölgy használata emelkedik, utóbbi intenzívebb emelkedésében a csírázásnak indult makkok játszatnak fontos szerepet. A használatnál tapasztalt kép tovább árnyalható az állományok preferenciájának vizsgálatával. A tölgy állományok esetében ez azt jelenti, hogy május, november, december hónapokon kívül az év többi szakaszában nem preferálta ezeket az élőhelyeket, ez alátámasztja azt, hogy november - december hónapokban a lehullott makk, májusban pedig a csírázó makk teszi vonzóvá a tölgyeseket. Az égeresek esetében csak a júniustól-decemberig tartó időszakban volt megfigyelhető a preferencia, mértéke szeptembertől csökkenést mutat. Ezt magyarázza a már korábban említett búvóhelynek alkalmas takarás visszaszorulása. Hasonló preferencia volt kimutatható éger esetében gímszarvasnál is (TARI ET AL., 2009). A beállóhelyek vizsgálatának eredményei a tölgy esetében már a 0-10 éves korosztály esetében magas használatot mutattak ki, ami fokozódott a 10-20 éves korosztályban. Az éger esetében a 10-20 éves illetve az 50-60 éves korosztályt használta intenzívebben, ami a takarás megfelelő voltára utal.
Irodalomjegyzék
BIRKAN, M., SERRE, D., SKIBIESKI, S. AND PELARD, E., (1992): Spring-summer home range, habitat use and survival of grey partridge (Perdix perdix) in a semi-open habitat, Game and Wildlife, 9:431-442
FOURNIER CHAMBRILLON, C., MAILLARD, D. AND FOURNIER, P (1995): Diet of wild boar (Sus scrofa L.) inhabiting the montpellier garrigue, IBEX J. M. E. 3:174-179
HERRERO, J., IRIZAR, I., LASKURAIN, N. A., GARCIA-SERRANO, A. AND GARCIA-GONZALEZ, R. (2005): Fruit and roots: wild boar foods during the cold season int he southwestern Pyrenees, Ital. J. Zool. ,72:49-52
HERRERO, J., GARCIA-SERRANO, A., COUTO, S., ORTUNO, V. M. AND GARCIA-GONZALES, R. (2006): Diet of wild boar Sus scrofa L. and crop damage in an intensive agroecosystem, Eur. J. Wildl. Res. 52:245-250
HOWE, T.D., SINGER, FJ. AND ACKERMAN, B. (1981): Forage relationships of European wild boar invading northern hardwood forest. J Wildl Manage 45:748–754
D’HUART, J.P. (1991): Habitat utilization of old world wild pigs. In: Barrett RG, Spitz F (eds) Biology of suidae. IRGM, Grenoble, pp 30–48.
KEULING, O., STIER, N., AND ROTH, M. (2008): How does hunting influence activity and spatial usage in wild boar Sus scrofa L.? Eur J Wildl Res 54(4): 729-737.
MAILLARD, D. AND FOURNIER, P. (1995): Effect of shooting with hounds on home range size of wild boar (Sus scrofa L.) groups in Mediterranean habitat. IBEX J Mt Ecol 3:102–107
MASSEI, G., GENOV, P. V., STAINES, B. W. AND GORMAN, M. L. (1997): Factors influencing home range and activity of wild boar (Sus scrofa) in a Mediterranean coastal areaJournal of Zoology 242(3):411 - 423.
MCDONALD, D. (2001): The new emcyklopedia of mammals. Oxford University Press, Oxford,
NÁHLIK, A., HEFFENTRÄGER, G., PÓCZA, G., SÁNDOR, GY. AND TARI, T. (2014): Daytime movements of red deer disturbed by human activity, Proceedings of The 8th Interbational Deer Biology Congress & International Wildlife Management Symposium, p.57-59.
NÁHLIK, A., SÁNDOR, GY., HEFFENTRÄGER, G., PÓCZA, G. ÉS TARI, T. (2014): Előzetes eredmények a vaddisznó (Sus scrofa) mozgáskörzetének alakulásáról, NYME Erdőmérnöki Kar Tudományos Konferencia, Összefoglaló Kötet p. 107
TARI T., SÁNDOR GY. ÉS NÁHLIK A., (2009): A gímszarvas élőhely-preferenciája egy síkvidéki területen, NYME Erdőmérnöki Kar Tudományos Konferencia, Konferencia Kiadvány p. 256-261.
----------------------
A következő napokban az alábbi előadás-anyagokat is olvashatja portálunkon az Alföldi Erdőkért Egyesület Kutatói Napjáról:
- Gondolatok az innovációs eredmények gyakorlati bevezetéséről, mint stratégiáról az alföldi erdőgazdálkodásban – ELOLVASOM >>>
- Energetikai ültetvények az Alföldön - ELOLVASOM >>>
- Szarvasgomba – gazdálkodás és kutatás a NEFAG Zrt-nél - ELOLVASOM >>>
- Akác virágzásbiológiai vizsgálatok Alföldi erdőgazdaságoknál - ELOLVASOM >>>
- Kedvezőtlen termőhelyi adottságú kocsányos tölgy állomány lékes felújításának állapotváltozása - ELOLVASOM >>>
- A vaddisznó területhasználata és aktivitása egy síkvidéki élőhelyen -
- Hagyományos és új agroerdészeti technológiák lehetséges szerepe az Alföld klímaérzékenységének mérséklésében - ELOLVASOM >>>
- Kocsányos tölgy erdőfelújítási technológiák összehasonlítása a Kisalföldi Erdőgazdaság területén lévő Rábakecöl 6/G erdőrészletben - ELOLVASOM >>>
- Az Alföldi Erdőkért Egyesület erdészeti részvénytársasági tagjainak átláthatósága - ELOLVASOM >>>
- Az erdők hatása a sófelhalmozódásra sekély talajvízű alföldi területeken - ELOLVASOM >>>
- Harveszterek terjedése Magyarországon - ELOLVASOM >>>
- A bálványfa (Ailanthus altissima) faipari és energetikai célú alkalmazhatósága - ELOLVASOM >>>
- Környezetkímélő és költséghatékony agroerdészeti termesztési rendszerek, mint a jövő földhasználati lehetőségei - ELOLVASOM >>>
- Király dió és magas kőris elegyes növekedésdinamikai elemzése az évgyűrűk alapján - ELOLVASOM >>>
- Talajtömörödöttség mérésére alapozott termőhely-értékelés tapasztalatai a Nyírségben - ELOLVASOM >>>
- Szélsőséges termőhelyi jellemzők hatása kocsányos tölgy erdőállomány talajnedvességére lékes felújítása során - ELOLVASOM >>>
- Akác kutatások a Faanyagtudományi Intézetben - ELOLVASOM >>>
- Rövid vágásfordulójú energetikai faültetvények technológiai sajátosságai - ELOLVASOM >>>
- Numerikus analízis alkalmazása forgó szerszám fejlesztéséhez - ELOLVASOM >>>
(Forrás: aee.hu - Engedéllyel közzétéve: Erdő-Mező Online – www.erdo-mezo.hu)
Hasonlóan megnövelt aktivitással reagált a gímszarvas a nem állandóan fennálló, intenzív zavaró hatásokra (NÁHLIK ET AL., 2014). Az erdei élőhely-használat vizsgálata során tapasztaltak a vaddisznó kiváló alkalmazkodó képességét támasztják alá (D’HUART, 1991).
Élőhely-használata során törekedett a számára legoptimálisabb faállományok kiválasztására, amelyek táplálkozás és búvóhely szempontjából is megfelelőek. A vizsgálati területen két fafaj állományait használta intenzíven, ezek a kocsányos tölgy és a mézgás éger voltak. A használat havi alakulása jól szemlélteti vaddisznó arra adott reakcióját, hogy a két fafaj állományai által nyújtott táplálék- és búvóhely kínálatban változások történnek. A mézgás éger, magas aljnövényzetével, elsősorban nappali búvóhelyet nyújt a vaddisznó számára, ezt támasztják alá a beállóhelyek használatának elemzésekor szerzett eredményeink. A nedves-vizes élőhely jó dagonyázási lehetőséget jelent, illetve táplálkozó helyként is szerepe lehet, a magas puhatestű táplálék kínálatból kifolyólag, ami kedvelt tápláléka a vaddisznónak (HOWE ET AL. 1981).
A fiatal tölgy állományok ugyanakkor búvóhelyet jelentenek, míg az idősebb állományok makktermésük révén töltenek be fontos szerepet a vaddisznó táplálkozásában, mivel a természetes táplálékok közül ez a legpreferáltabb forrás (FOURNIER-CHAMBRILLON ET AL., 1995). Vizsgálatunkban a tölgy állományok legalacsonyabb használati értéke augusztusban volt megfigyelhető, majd folyamatos emelkedés mellett, decemberben érte el a csúcsot, ezzel párhuzamosan viszont csökkent az égeresek használata. Az augusztus december periódus kezdetén a mezőgazdasági kultúrák nyújtotta táplálék kedveltsége (HERRERO ET AL. 2006) és az égeresek dús cserjeszintje okozhatja a tölgyek alacsony használatát. Majd az ősz során az égeresek takarásának csökkenésével és a mezőgazdasági kultúrák visszaszorulásával párhuzamosan növekedni kezd a tölgyesek használata. A magas novemberi-decemberi tölgy használatban elsősorban a táplálék kínálat játszhat szerepet, ami januárra visszaesik. Ez köszönhető a makk fogyásának, a hótakarónak és a fagyott talajnak, ugyan is a fagyott föld miatt csökken a földből felvett táplálékok aránya a táplálékban (HERRERO ET. AL, 2005). Az égeresek használatának márciusig folytatódó csökkenésével a tölgyesek használata emelkedik, ez az égeresek nyújtotta takarás további csökkenésével hozható összefüggésbe. A vegetáció megindulásával áprilistól kezdődően mind az éger, mind pedig a tölgy használata emelkedik, utóbbi intenzívebb emelkedésében a csírázásnak indult makkok játszatnak fontos szerepet. A használatnál tapasztalt kép tovább árnyalható az állományok preferenciájának vizsgálatával. A tölgy állományok esetében ez azt jelenti, hogy május, november, december hónapokon kívül az év többi szakaszában nem preferálta ezeket az élőhelyeket, ez alátámasztja azt, hogy november - december hónapokban a lehullott makk, májusban pedig a csírázó makk teszi vonzóvá a tölgyeseket. Az égeresek esetében csak a júniustól-decemberig tartó időszakban volt megfigyelhető a preferencia, mértéke szeptembertől csökkenést mutat. Ezt magyarázza a már korábban említett búvóhelynek alkalmas takarás visszaszorulása. Hasonló preferencia volt kimutatható éger esetében gímszarvasnál is (TARI ET AL., 2009). A beállóhelyek vizsgálatának eredményei a tölgy esetében már a 0-10 éves korosztály esetében magas használatot mutattak ki, ami fokozódott a 10-20 éves korosztályban. Az éger esetében a 10-20 éves illetve az 50-60 éves korosztályt használta intenzívebben, ami a takarás megfelelő voltára utal.
Irodalomjegyzék
BIRKAN, M., SERRE, D., SKIBIESKI, S. AND PELARD, E., (1992): Spring-summer home range, habitat use and survival of grey partridge (Perdix perdix) in a semi-open habitat, Game and Wildlife, 9:431-442
FOURNIER CHAMBRILLON, C., MAILLARD, D. AND FOURNIER, P (1995): Diet of wild boar (Sus scrofa L.) inhabiting the montpellier garrigue, IBEX J. M. E. 3:174-179
HERRERO, J., IRIZAR, I., LASKURAIN, N. A., GARCIA-SERRANO, A. AND GARCIA-GONZALEZ, R. (2005): Fruit and roots: wild boar foods during the cold season int he southwestern Pyrenees, Ital. J. Zool. ,72:49-52
HERRERO, J., GARCIA-SERRANO, A., COUTO, S., ORTUNO, V. M. AND GARCIA-GONZALES, R. (2006): Diet of wild boar Sus scrofa L. and crop damage in an intensive agroecosystem, Eur. J. Wildl. Res. 52:245-250
HOWE, T.D., SINGER, FJ. AND ACKERMAN, B. (1981): Forage relationships of European wild boar invading northern hardwood forest. J Wildl Manage 45:748–754
D’HUART, J.P. (1991): Habitat utilization of old world wild pigs. In: Barrett RG, Spitz F (eds) Biology of suidae. IRGM, Grenoble, pp 30–48.
KEULING, O., STIER, N., AND ROTH, M. (2008): How does hunting influence activity and spatial usage in wild boar Sus scrofa L.? Eur J Wildl Res 54(4): 729-737.
MAILLARD, D. AND FOURNIER, P. (1995): Effect of shooting with hounds on home range size of wild boar (Sus scrofa L.) groups in Mediterranean habitat. IBEX J Mt Ecol 3:102–107
MASSEI, G., GENOV, P. V., STAINES, B. W. AND GORMAN, M. L. (1997): Factors influencing home range and activity of wild boar (Sus scrofa) in a Mediterranean coastal areaJournal of Zoology 242(3):411 - 423.
MCDONALD, D. (2001): The new emcyklopedia of mammals. Oxford University Press, Oxford,
NÁHLIK, A., HEFFENTRÄGER, G., PÓCZA, G., SÁNDOR, GY. AND TARI, T. (2014): Daytime movements of red deer disturbed by human activity, Proceedings of The 8th Interbational Deer Biology Congress & International Wildlife Management Symposium, p.57-59.
NÁHLIK, A., SÁNDOR, GY., HEFFENTRÄGER, G., PÓCZA, G. ÉS TARI, T. (2014): Előzetes eredmények a vaddisznó (Sus scrofa) mozgáskörzetének alakulásáról, NYME Erdőmérnöki Kar Tudományos Konferencia, Összefoglaló Kötet p. 107
TARI T., SÁNDOR GY. ÉS NÁHLIK A., (2009): A gímszarvas élőhely-preferenciája egy síkvidéki területen, NYME Erdőmérnöki Kar Tudományos Konferencia, Konferencia Kiadvány p. 256-261.
----------------------
A következő napokban az alábbi előadás-anyagokat is olvashatja portálunkon az Alföldi Erdőkért Egyesület Kutatói Napjáról:
- Gondolatok az innovációs eredmények gyakorlati bevezetéséről, mint stratégiáról az alföldi erdőgazdálkodásban – ELOLVASOM >>>
- Energetikai ültetvények az Alföldön - ELOLVASOM >>>
- Szarvasgomba – gazdálkodás és kutatás a NEFAG Zrt-nél - ELOLVASOM >>>
- Akác virágzásbiológiai vizsgálatok Alföldi erdőgazdaságoknál - ELOLVASOM >>>
- Kedvezőtlen termőhelyi adottságú kocsányos tölgy állomány lékes felújításának állapotváltozása - ELOLVASOM >>>
- A vaddisznó területhasználata és aktivitása egy síkvidéki élőhelyen -
- Hagyományos és új agroerdészeti technológiák lehetséges szerepe az Alföld klímaérzékenységének mérséklésében - ELOLVASOM >>>
- Kocsányos tölgy erdőfelújítási technológiák összehasonlítása a Kisalföldi Erdőgazdaság területén lévő Rábakecöl 6/G erdőrészletben - ELOLVASOM >>>
- Az Alföldi Erdőkért Egyesület erdészeti részvénytársasági tagjainak átláthatósága - ELOLVASOM >>>
- Az erdők hatása a sófelhalmozódásra sekély talajvízű alföldi területeken - ELOLVASOM >>>
- Harveszterek terjedése Magyarországon - ELOLVASOM >>>
- A bálványfa (Ailanthus altissima) faipari és energetikai célú alkalmazhatósága - ELOLVASOM >>>
- Környezetkímélő és költséghatékony agroerdészeti termesztési rendszerek, mint a jövő földhasználati lehetőségei - ELOLVASOM >>>
- Király dió és magas kőris elegyes növekedésdinamikai elemzése az évgyűrűk alapján - ELOLVASOM >>>
- Talajtömörödöttség mérésére alapozott termőhely-értékelés tapasztalatai a Nyírségben - ELOLVASOM >>>
- Szélsőséges termőhelyi jellemzők hatása kocsányos tölgy erdőállomány talajnedvességére lékes felújítása során - ELOLVASOM >>>
- Akác kutatások a Faanyagtudományi Intézetben - ELOLVASOM >>>
- Rövid vágásfordulójú energetikai faültetvények technológiai sajátosságai - ELOLVASOM >>>
- Numerikus analízis alkalmazása forgó szerszám fejlesztéséhez - ELOLVASOM >>>
(Forrás: aee.hu - Engedéllyel közzétéve: Erdő-Mező Online – www.erdo-mezo.hu)
















