



2026. augusztus 8. szombat,
László napja
László napja
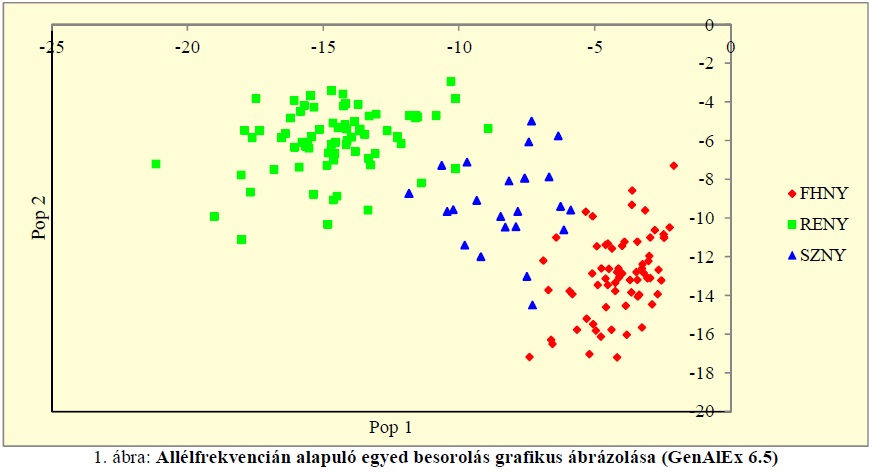 Az ábrán jól látható, hogy a hibrid szürke nyár egyedek a két alapfaj egyedei alkotta pontfelhő között helyezkednek el a genetikai adatok alapján. A hibrid egyedek körének pontosítását követően elvégeztük a vizsgált populációk genetikai távolságának meghatározását. Ennek érdekében a populációgenetikai program segítségével kiszámítottuk az egyes populációk Nei-féle genetikai távolságát. Az elkészített genetikai távolság mátrix alapján a populációk kapcsolatát Statistica 6.0 program segítségével dendrogramon ábrázoltuk (2. ábra). A klaszterek kialakítása a Statistica 6.0 szoftver Complete linkage módszerével történt, amely az egymástól legtávolabbi genotípusok összevetésén alapszik (Podani 1997).
Az ábrán jól látható, hogy a hibrid szürke nyár egyedek a két alapfaj egyedei alkotta pontfelhő között helyezkednek el a genetikai adatok alapján. A hibrid egyedek körének pontosítását követően elvégeztük a vizsgált populációk genetikai távolságának meghatározását. Ennek érdekében a populációgenetikai program segítségével kiszámítottuk az egyes populációk Nei-féle genetikai távolságát. Az elkészített genetikai távolság mátrix alapján a populációk kapcsolatát Statistica 6.0 program segítségével dendrogramon ábrázoltuk (2. ábra). A klaszterek kialakítása a Statistica 6.0 szoftver Complete linkage módszerével történt, amely az egymástól legtávolabbi genotípusok összevetésén alapszik (Podani 1997).
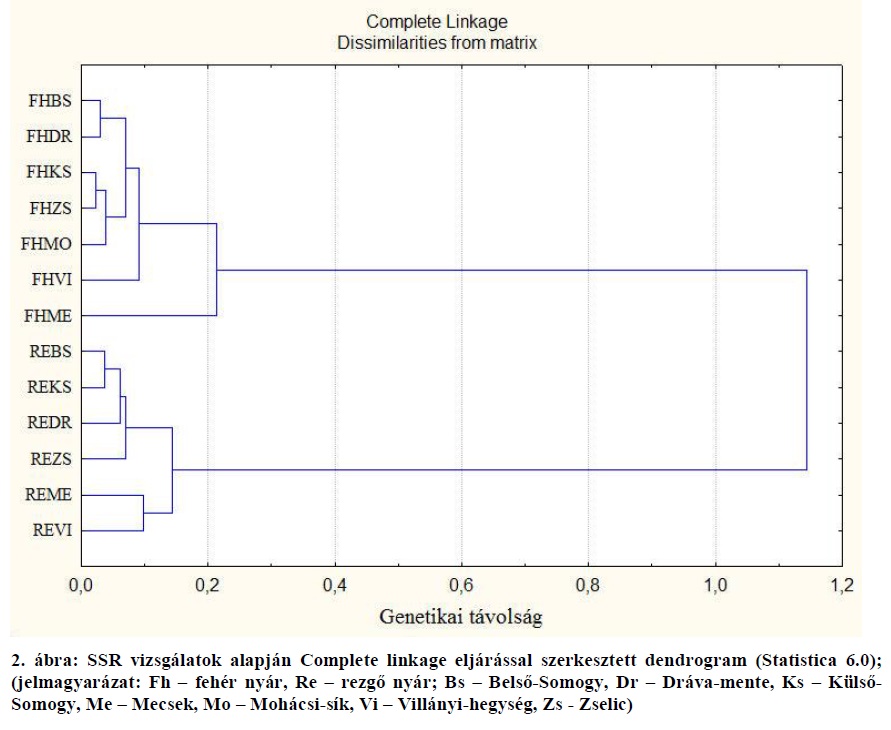 A dendrogram alapján általánosságban megállapítható, hogy a földrajzilag egymáshoz közel fekvő populációk genetikai távolsága kisebb, mint a földrajzilag egymástól távol esőké. Ezt támasztja alá a fehér nyár esetében a belső-somogyi és dráva-menti, valamint a külső-somogyi és zselici populációk, továbbá a rezgő nyár esetében a belső- és külső-somogyi, populációk kis genetikai távolsága. A somogyi rezgő nyár populációk genetikai távolsága a dráva-menti és zselici populációkhoz viszonyítva sem jelentős. Ugyanakkor mindkét faj esetében megállapítható, hogy a Villányi-hegységben és a Mecsekben mintázott egyedek genetikailag távolabb esnek a szomszédos populációktól (ezt ugyanakkor a kis egyedszám is magyarázhatja, hiszen a tájegységek populációit a fehér nyár esetében csupán két-két egyed, a rezgő nyár esetében sorrendben 7 és 9 egyed képviseli).
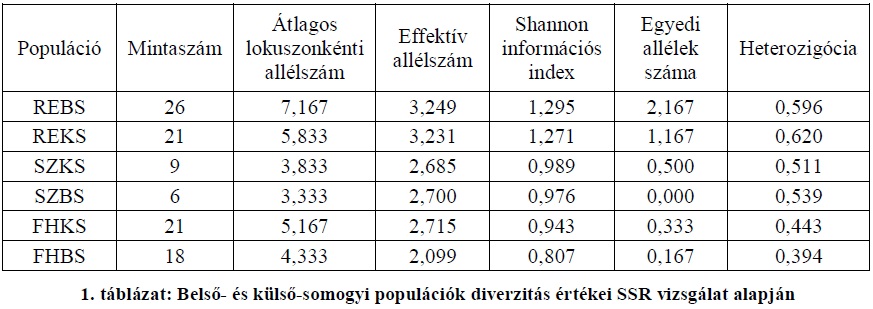
A somogyi populációk SSR vizsgálatok során nyert diverzitás értékeit külön ábrázoljuk (1. táblázat). A táblázatban az allélszámot, valamint az allélgyakoriságokat figyelembe vevő Shannon információs index értékek alapján rangsoroltuk az egyes populációkat. A táblázat alapján jól látható, hogy Shannon-index értékeik alapján a rezgő nyár populációk mutatják a legmagasabb genetikai diverzitást. Bár az átlagos lokuszonkénti allélszámban és egyedi allélek számában a belső-somogyi rezgő nyár populáció kiemelkedik a többi populáció közül, a várt heterozigócia esetében a külső-somogyi rezgő nyár populáció mutat magasabb értéket. A Shannon-index alapján értékelt diverzitás tekintetében a szürke nyár populációk a két alapfaj populációi között helyezkednek el, Külső-Somogy, Belső-Somogy sorrendben. A vizsgált fehér nyár populációk mérsékelt diverzitást mutattak a másik két faj populációihoz képest.
A genetikai variancia megoszlásának meghatározásához molekuláris genetikai varianciaanalízist (AMOVA-teszt) végeztünk a GenAlEx 6.5 programmal, a somogyi populációkban mintázott egyedek bevonásával. A varianciaanalízis eredményeképpen megállapítható, hogy a teljes genetikai varianciához legnagyobb mértékben az egyedszintű variancia járul hozzá (74%). Azt követi a populációk közötti (22%), majd az egyedek közötti variancia (4%).
Összefoglalás
A Közép- és Dél-Dunántúl 7 tájegységében gyűjtött összesen 169 őshonos Leuce nyár egyeden 6 markerrel végzett mikroszatellit (SSR) analízis alkalmasnak bizonyult az alapfajok (fehér nyár, rezgő nyár), valamint a hibridegyedek (szürke nyár) laboratóriumi elkülönítésére. A szürke nyár a két alapfaj közötti, átmeneti genetikai mintázatot mutatott. A Nei-féle genetikai távolság alapján szerkesztett dendrogram a földrajzilag egymáshoz közel fekvő populációk tekintetében jelzett kisebb genetikai távolságot, ami a populációk közötti erőteljes génáramlásra hívja fel a figyelmet. A populációgenetikai szoftverrel számított diverzitás értékek a somogyi rezgő nyár populáció kiemelkedő változatosságát mutatták. Végezetül a somogyi populációkban mért molekuláris variancia forrását, vizsgálataink szerint, döntő mértékben az egyedszintű variancia határozza meg (74%).
Köszönetnyilvánítás
A mintagyűjtés alkalmával segítőink, vezetőink voltak a Duna-Dráva Nemzeti Park, a Mecseki Erdészeti ZRt., a SEFAG Erdészeti és Faipari ZRt. munkatársai, akiknek ezúton is szeretnénk kifejezni hálás köszönetünket fáradozásaikért. Kutatásunkat az OTKA 06321-es nyilvántartási számú pályázata támogatta.
Felhasznált irodalom
Bartha D. (2004): A magyarországi nyár (Populus L.) taxonok határozókulcsa és rövid jellemzése. Flora Pannonica 2(2): 93.
Hajósné Novák M. (1999): Genetikai variabilitás a növénynemesítésben. Mezőgazda Kiadó, Budapest, p. 44-48.
Koltay Gy.,Kopecky F. (1954): Őshonos nyáraink leromlott öröklöttségének megjavítása. Erdészeti Kutatások, 2: 65-86.
NÉBIH Erdészeti Igazgatóság (2013): Erdővagyon, erdőgazdálkodás Magyarországon. http://www.nebih.gov.hu/szakteruletek/szakteruletek/erdeszeti_igazgatosag/kozerdeku_adatok/adatok
Peakall, R., Smouse, P.E. (2012): GENALEX 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 28, 2537-2539.
Podani J. (1997): Bevezetés a többváltozós biológiai adatfeltárás rejtelmeibe. Scientia Kiadó. Budapest, p. 145.
StatSoft, Inc. (2001). STATISTICA for Windows [Computer program manual]. Tulsa, OK:
StatSoft, Inc., 2300 East 14th Street, Tulsa, OK 74104, phone: (918) 749-1119, fax: (918) 749-2217, email: info@statsoft.com, WEB: http://www.statsoft.com
-------
A következő napokban az alábbi előadás-anyagokat is olvashatja portálunkon az Alföldi Erdőkért Egyesület Kutatói Napjáról:
- A lakitelki erdők leírása, különös tekintettel az őshonosság tükrében - ELOLVASOM >>>
- Az őshonosság fogalmával kapcsolatos elvárások és kétségek - ELOLVASOM >>>
- A Két Víz közének védett és Natura 2000 erdei a változó környezeti viszonyok mellett - ELOLVASOM >>>
- A Szatmár-Beregi kocsányos tölgyesek erdőgazdálkodási tapasztalatai - ELOLVASOM >>>
- Fehérnyár klónok teljesítményvizsgálata alföldi száraz, homoki termőhelyen - ELOLVASOM >>>
- Őshonos nyárfajok genetikai változatossága a Dunántúlon -------
- Szolnoki árapasztó csatornában végzett beavatkozások árvízre gyakorolt hatásai - ELOLVASOM >>>
- Új hazai, ERTI szelekciójú fűz klón teljesítmény vizsgálata szabadalmi bejelentés megalapozására - ELOLVASOM >>>
- Szelektált akác származások virágzásbiológiai vizsgálata Tiszántúl száraz homoki termőhelyen - ELOLVASOM >>>
- Kocsányos tölgy állomány talajnedvességének változása lékes felújítás során - ELOLVASOM >>>
- Hazai kocsánytalan tölgy állományok faanyag-minőségi kérdései- ELOLVASOM >>>
- Kisalföldi nemesnyár hibridek szárítása – ELOLVASOM >>>
- A magyarországi erdők energetikai célra hasznosítható faanyaga – ELOLVASOM >>>
- Energetikai ültetvények égési jellemzőinek vizsgálata – ELOLVASOM >>>
- A biomassza termesztés és feldolgozás függősége a klímaváltozástól - ELOLVASOM >>>
- Rövid vágásfordulójú energetikai ültetvények lombtömegének vizsgálata, szénkörforgalomban betöltött szerepe – ELOLVASOM >>>
- Forvarderek alkalmazási lehetőségei napjainkban - ELOLVASOM >>>
- Talajművelő szerszám végeselem modellezése ELOLVASOM >>>
- Repceszárból előállított pellet hasznosításának ökoenergetikai kérdései – ELOLVASOM >>>
(Forrás: aee.hu - Engedéllyel közzétéve: Erdő-Mező Online - www.erdo-mezo.hu)
A dendrogram alapján általánosságban megállapítható, hogy a földrajzilag egymáshoz közel fekvő populációk genetikai távolsága kisebb, mint a földrajzilag egymástól távol esőké. Ezt támasztja alá a fehér nyár esetében a belső-somogyi és dráva-menti, valamint a külső-somogyi és zselici populációk, továbbá a rezgő nyár esetében a belső- és külső-somogyi, populációk kis genetikai távolsága. A somogyi rezgő nyár populációk genetikai távolsága a dráva-menti és zselici populációkhoz viszonyítva sem jelentős. Ugyanakkor mindkét faj esetében megállapítható, hogy a Villányi-hegységben és a Mecsekben mintázott egyedek genetikailag távolabb esnek a szomszédos populációktól (ezt ugyanakkor a kis egyedszám is magyarázhatja, hiszen a tájegységek populációit a fehér nyár esetében csupán két-két egyed, a rezgő nyár esetében sorrendben 7 és 9 egyed képviseli).
A somogyi populációk SSR vizsgálatok során nyert diverzitás értékeit külön ábrázoljuk (1. táblázat). A táblázatban az allélszámot, valamint az allélgyakoriságokat figyelembe vevő Shannon információs index értékek alapján rangsoroltuk az egyes populációkat. A táblázat alapján jól látható, hogy Shannon-index értékeik alapján a rezgő nyár populációk mutatják a legmagasabb genetikai diverzitást. Bár az átlagos lokuszonkénti allélszámban és egyedi allélek számában a belső-somogyi rezgő nyár populáció kiemelkedik a többi populáció közül, a várt heterozigócia esetében a külső-somogyi rezgő nyár populáció mutat magasabb értéket. A Shannon-index alapján értékelt diverzitás tekintetében a szürke nyár populációk a két alapfaj populációi között helyezkednek el, Külső-Somogy, Belső-Somogy sorrendben. A vizsgált fehér nyár populációk mérsékelt diverzitást mutattak a másik két faj populációihoz képest.
A genetikai variancia megoszlásának meghatározásához molekuláris genetikai varianciaanalízist (AMOVA-teszt) végeztünk a GenAlEx 6.5 programmal, a somogyi populációkban mintázott egyedek bevonásával. A varianciaanalízis eredményeképpen megállapítható, hogy a teljes genetikai varianciához legnagyobb mértékben az egyedszintű variancia járul hozzá (74%). Azt követi a populációk közötti (22%), majd az egyedek közötti variancia (4%).
Összefoglalás
A Közép- és Dél-Dunántúl 7 tájegységében gyűjtött összesen 169 őshonos Leuce nyár egyeden 6 markerrel végzett mikroszatellit (SSR) analízis alkalmasnak bizonyult az alapfajok (fehér nyár, rezgő nyár), valamint a hibridegyedek (szürke nyár) laboratóriumi elkülönítésére. A szürke nyár a két alapfaj közötti, átmeneti genetikai mintázatot mutatott. A Nei-féle genetikai távolság alapján szerkesztett dendrogram a földrajzilag egymáshoz közel fekvő populációk tekintetében jelzett kisebb genetikai távolságot, ami a populációk közötti erőteljes génáramlásra hívja fel a figyelmet. A populációgenetikai szoftverrel számított diverzitás értékek a somogyi rezgő nyár populáció kiemelkedő változatosságát mutatták. Végezetül a somogyi populációkban mért molekuláris variancia forrását, vizsgálataink szerint, döntő mértékben az egyedszintű variancia határozza meg (74%).
Köszönetnyilvánítás
A mintagyűjtés alkalmával segítőink, vezetőink voltak a Duna-Dráva Nemzeti Park, a Mecseki Erdészeti ZRt., a SEFAG Erdészeti és Faipari ZRt. munkatársai, akiknek ezúton is szeretnénk kifejezni hálás köszönetünket fáradozásaikért. Kutatásunkat az OTKA 06321-es nyilvántartási számú pályázata támogatta.
Felhasznált irodalom
Bartha D. (2004): A magyarországi nyár (Populus L.) taxonok határozókulcsa és rövid jellemzése. Flora Pannonica 2(2): 93.
Hajósné Novák M. (1999): Genetikai variabilitás a növénynemesítésben. Mezőgazda Kiadó, Budapest, p. 44-48.
Koltay Gy.,Kopecky F. (1954): Őshonos nyáraink leromlott öröklöttségének megjavítása. Erdészeti Kutatások, 2: 65-86.
NÉBIH Erdészeti Igazgatóság (2013): Erdővagyon, erdőgazdálkodás Magyarországon. http://www.nebih.gov.hu/szakteruletek/szakteruletek/erdeszeti_igazgatosag/kozerdeku_adatok/adatok
Peakall, R., Smouse, P.E. (2012): GENALEX 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 28, 2537-2539.
Podani J. (1997): Bevezetés a többváltozós biológiai adatfeltárás rejtelmeibe. Scientia Kiadó. Budapest, p. 145.
StatSoft, Inc. (2001). STATISTICA for Windows [Computer program manual]. Tulsa, OK:
StatSoft, Inc., 2300 East 14th Street, Tulsa, OK 74104, phone: (918) 749-1119, fax: (918) 749-2217, email: info@statsoft.com, WEB: http://www.statsoft.com
-------
A következő napokban az alábbi előadás-anyagokat is olvashatja portálunkon az Alföldi Erdőkért Egyesület Kutatói Napjáról:
- A lakitelki erdők leírása, különös tekintettel az őshonosság tükrében - ELOLVASOM >>>
- Az őshonosság fogalmával kapcsolatos elvárások és kétségek - ELOLVASOM >>>
- A Két Víz közének védett és Natura 2000 erdei a változó környezeti viszonyok mellett - ELOLVASOM >>>
- A Szatmár-Beregi kocsányos tölgyesek erdőgazdálkodási tapasztalatai - ELOLVASOM >>>
- Fehérnyár klónok teljesítményvizsgálata alföldi száraz, homoki termőhelyen - ELOLVASOM >>>
- Őshonos nyárfajok genetikai változatossága a Dunántúlon -------
- Szolnoki árapasztó csatornában végzett beavatkozások árvízre gyakorolt hatásai - ELOLVASOM >>>
- Új hazai, ERTI szelekciójú fűz klón teljesítmény vizsgálata szabadalmi bejelentés megalapozására - ELOLVASOM >>>
- Szelektált akác származások virágzásbiológiai vizsgálata Tiszántúl száraz homoki termőhelyen - ELOLVASOM >>>
- Kocsányos tölgy állomány talajnedvességének változása lékes felújítás során - ELOLVASOM >>>
- Hazai kocsánytalan tölgy állományok faanyag-minőségi kérdései- ELOLVASOM >>>
- Kisalföldi nemesnyár hibridek szárítása – ELOLVASOM >>>
- A magyarországi erdők energetikai célra hasznosítható faanyaga – ELOLVASOM >>>
- Energetikai ültetvények égési jellemzőinek vizsgálata – ELOLVASOM >>>
- A biomassza termesztés és feldolgozás függősége a klímaváltozástól - ELOLVASOM >>>
- Rövid vágásfordulójú energetikai ültetvények lombtömegének vizsgálata, szénkörforgalomban betöltött szerepe – ELOLVASOM >>>
- Forvarderek alkalmazási lehetőségei napjainkban - ELOLVASOM >>>
- Talajművelő szerszám végeselem modellezése ELOLVASOM >>>
- Repceszárból előállított pellet hasznosításának ökoenergetikai kérdései – ELOLVASOM >>>
(Forrás: aee.hu - Engedéllyel közzétéve: Erdő-Mező Online - www.erdo-mezo.hu)
















